For an up-to-date list of publications, see Google Scholar.
* and + denote joint authorship.
2026
-
Phasic dopamine drives conditioned responding beyond its role in learning
bioRxiv (2026) link pdf
Animals learn to respond to neutral stimuli paired with rewards. Temporal Difference (TD) learning suggests animals learn to predict future reward value, updated via reward prediction errors (RPE) signaled by dopamine neurons. Traditionally, conditioned responses are thought to reflect value. However, here we use data analysis and modeling to argue that conditioned responding reflects dopamine RPEs. This suggests that in addition to its role in learning, dopamine also directly influences conditioned responding.
2025
-
A theory of brain-computer interface learning via low-dimensional control
eLife (2025) link pdf
Primate motor systems show remarkable flexibility, exemplified by their ability to learn brain-computer interface (BCI) control---a novel motor behavior. And yet some BCI tasks are harder to learn than others. We propose a theory where learning occurs via a re-aiming strategy in a low-dimensional subspace of task-relevant inputs. This theory unifies findings across BCI tasks, makes testable predictions, and links learning to biological constraints on neural activity.
-
Prospective contingency explains behavior and dopamine signals during associative learning
Nature Neuroscience (2025) link pdf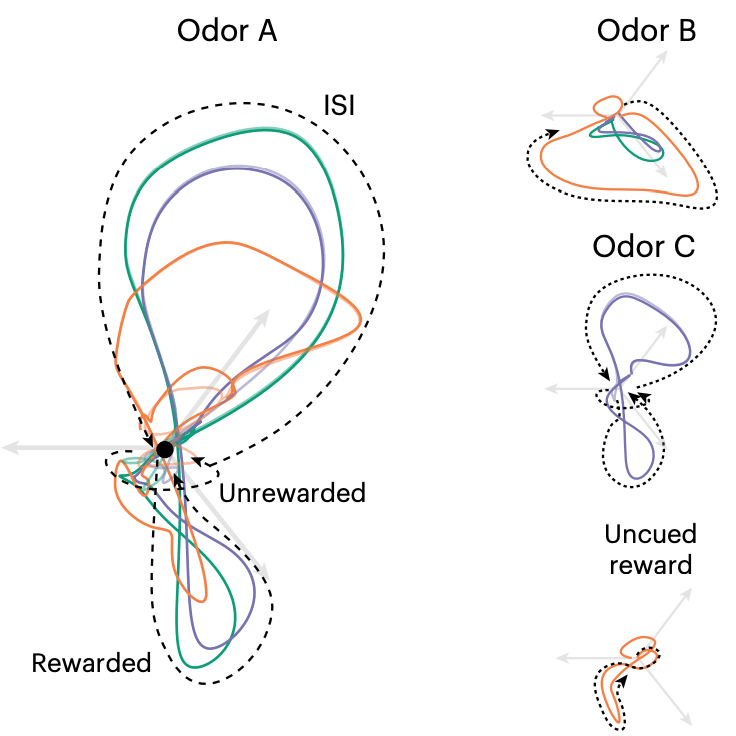
Associative learning depends on contingency, the degree to which a stimulus predicts an outcome. Curiously, we found that both anticipatory licking and dopamine responses to a conditioned stimulus decreased when animals received uncued rewards, but remained unchanged when animals instead received additional cued rewards. This finding cannot be explained by traditional accounts of contingency, but we show that it can be explained by temporal difference models with an appropriate state representation.
2024
-
Learning leaves a memory trace in motor cortex
Current Biology (2024) link pdf
How do we learn new behaviors without disrupting previously learned ones? Using a brain-computer interface (BCI) paradigm, we found that learning a new task altered the neural activity used to perform familiar tasks. This "memory trace" did not interfere with the performance of familiar tasks, suggesting a possible mechanism for how we learn new behaviors without impacting previously learned ones.
2023
-
Emergence of belief-like representations through reinforcement learning
PLOS Computational Biology (2023) link code pdf
Animals are thought to predict rewards using reinforcement learning (RL). In environments with hidden states, animals may require "beliefs," or probabilistic estimates of the hidden states. We show that such belief-like representations emerge in recurrent neural networks (RNN) trained to perform RL in environments with hidden states.
2021
-
How learning unfolds in the brain: toward an optimization view
Neuron (2021) link pdf
In this perspective, we consider the idea that learning in the brain can be described in terms of optimization, similar to learning in artificial neural networks (ANNs). We highlight three key features of how neural population changes with learning that differ from ANNs, suggesting refinements to this optimization view.
-
Learning is shaped by abrupt changes in neural engagement
Nature Neuroscience (2021) link code pdf
We identified large fluctuations in neural population activity in motor cortex (M1) indicative of arousal-like internal state changes. These changes in neural activity helped to explain why animals learned some tasks more quickly than others.
2020
-
Intracortical Brain-Machine Interfaces
Neural Engineering, 3rd Edition (2020) link pdf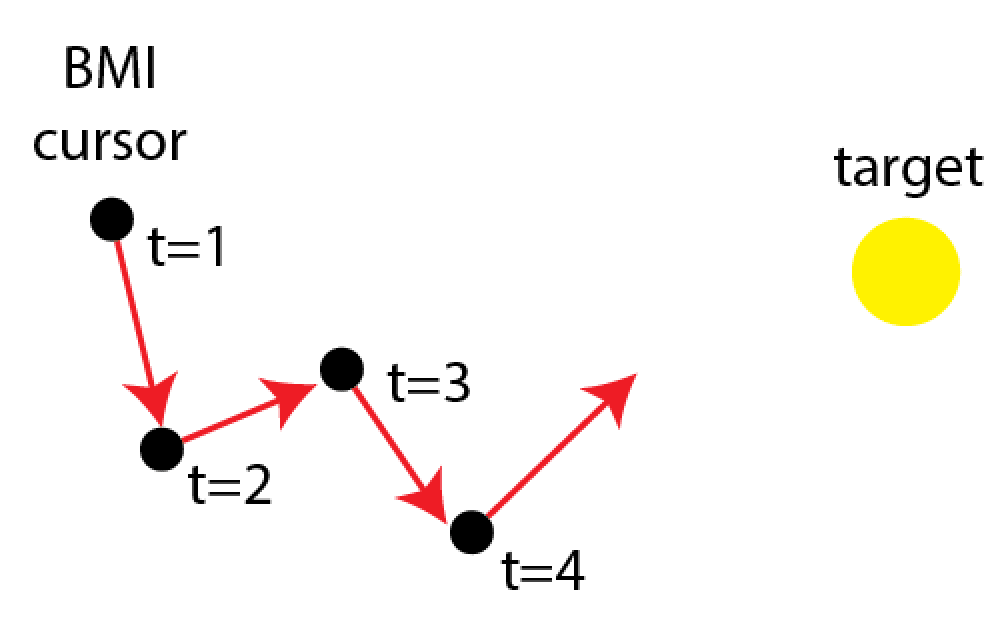
A brain–machine interface (BMI) directly connects the brain to the external world, translating a user's internal motor commands into action. In this chapter, we discuss the four basic components of an intracortical BMI: an intracortical neural recording, a decoding algorithm, an output device, and sensory feedback.
2019
-
New neural activity patterns emerge with long-term learning
Proceedings of the National Academy of Sciences (2019) link pdf
We establish that new neural activity patterns emerge with learning, providing evidence that the formation of new patterns of neural population activity can underlie the learning of new skills.
2018
-
Constraints on neural redundancy
eLife (2018) link code pdf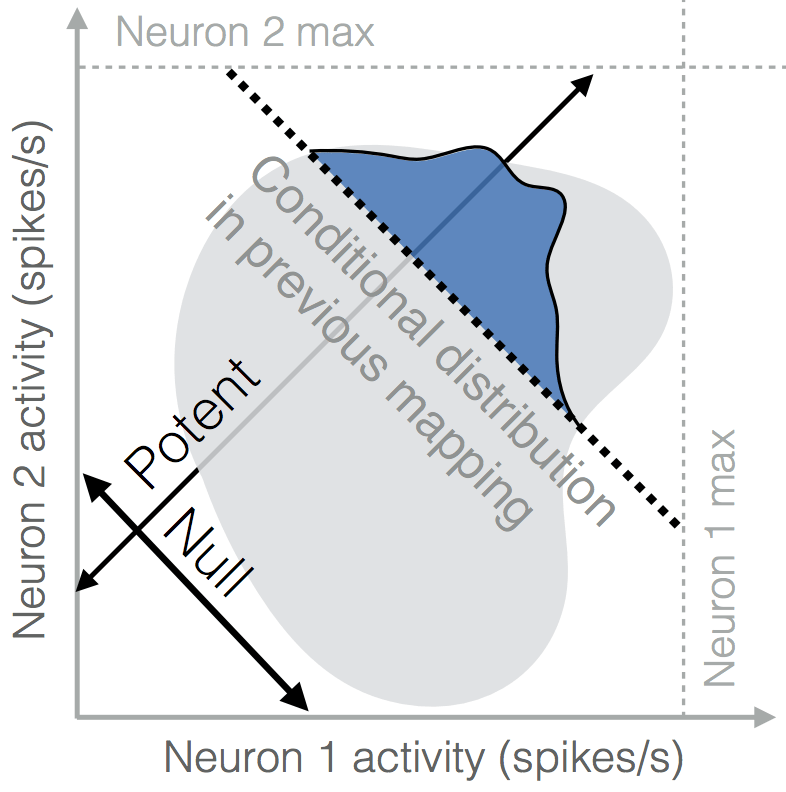
Millions of neurons in the brain control the activity of tens of muscles in the arm, meaning neural activity is redundant. We compared various hypotheses for how the brain deals with this redundancy by recording in primary motor cortex while subjects performed a brain-computer interface task.
2017
-
A classifying variational autoencoder with application to polyphonic music generation
arXiv (2017) link code pdf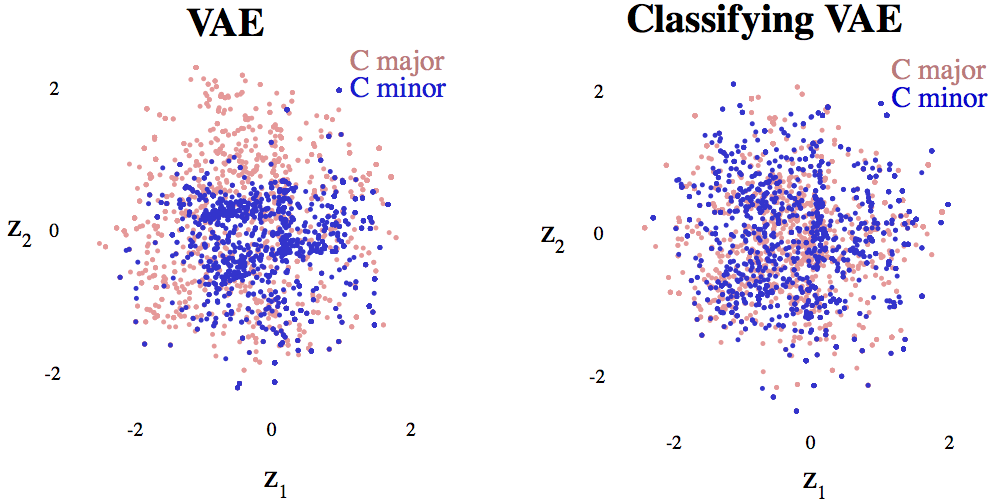
We augment a neural network known as a variational autoencoder (VAE) to classify the observed data while also learning its latent representation. We show that when this network is combined with an LSTM and used to generate music, the network plays fewer incorrect notes than a standard VAE+LSTM.
2015
-
A distinct mechanism of temporal integration for motion through depth
Journal of Neuroscience (2015) link pdf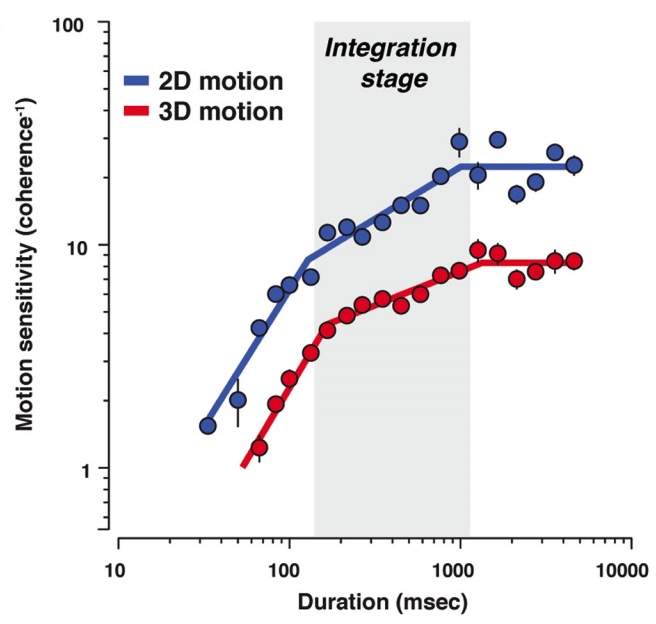
We compare the time-varying improvements in sensitivity during motion discrimination tasks in 2D and 3D, and find that the two are remarkably similar, however with a lower signal-to-noise ratio in 3D.
2013
-
Signal multiplexing and single-neuron computations in lateral intraparietal area during decision-making
Journal of Neuroscience (2013) link pdf
We show that cells in the lateral intraparietal area (LIP) have firing activity that simultaneously carries decision signals and decision-irrelevant sensory signals. We conclude that LIP cells show a broader range of response motifs than previously considered.